Extended Observation Descriptions
Cytological Observations on Desmids
Desmidiacea are always a feast for the eyes when observed under a microscope! The rich green, the symmetrical, often star-like shapes, the fine ornamentation on the cell walls delight the observers. Desmidiacea belong to the Zygnematophycea; this group of algae is at the foot of the phylogenetic tree of land plants in terms of evolutionary history and is therefore most closely related to them (Hess & al., 2022).
A Look into the World of Cell Organelles

Fig. 1: Nuclei with nucleoli of Closterium lunula and Micrasterias denticulata, including the winged and, in some species, finely lobed chloroplasts of Closterium striolatum, Netrium digitus var. lamellosum, Planotaenium interruptum, Xanthidium antilopaeum and Netrium oblongum.
If one wishes to observe organelles of desmids in the living state without vital (=sublethal) staining, their often beautifully structured chloroplasts are of course easy to observe, and the cell nuclei can often also be seen quite well, provided they are not masked by cell wall thickenings or overlying chloroplasts (Fig. 1).
The large, flat cells of the species Micrasterias rotata, Micrasterias denticulata, Micrasterias apiculata and Micrasterias thomasiana var. notata also allow the light microscopist an insight into the area of very small, low-contrast and therefore difficult to observe cell organelles such as dictyosomes and mitochondria (Fig. 2).

Fig. 2: Top row: Micrasterias rotata and M. denticulata. Bottom row: M. thomasiana var. notata and M. apiculata.
Scale bar indicates 100 µm.
Mitochondria are found in every plant or animal cell. They contain all the essential enzymes of the citric acid cycle and the respiratory chain, as well as fatty acid metabolism, along with those for the associated energy-producing phosphorylation processes. As the most important ATP suppliers, mitochondria are the centers for energy production from the respiratory processes in which the energy-rich carbon compounds are broken down. They usually have an elongated shape in the order of 0.2–1 µm by 2–8 µm, which corresponds to the size of bacteria.
Dictyosomes, also known as the Golgi apparatus, are centers for the synthesis of building materials for the cell. They appear as stacks of disc-shaped, flattened, membrane-bounded cavities or cisternae, whose diameter is usually 1–2 µm. Membrane-enclosed packages, the Golgi vesicles, are deposited at the edges of the cisternae. These are used to transport the Golgi secretions within the cell to their respective destinations. The dictyosomes of algae and plant cells primarily synthesize cell wall substances, acidic polysaccharides, pectins and hemicelluloses, but mucilage and essential oils are also produced. The fine silica plates for the tests of euglyphid testate amoebae are also products of dictyosomes, to name just another example from a completely different area of organisms (Fig. 3).

Fig. 3, from left: Assulina seminulum with its siliceous scales arranged like roof tiles; Cyphoderia ampulla, segment of a crushed specimen of the shell, whose rounded scales overlap so that the image resembles a chainmail; Tracheleuglypha dentata: the spreading of the silicate scales at the edge of the shell, induced by the light pressure of the cover glass, reveals how thin the silicate platelets are.
When euglyphid cells are about to divide, the silica scales produced in advance by the Golgi apparatus for the next generation can be seen lying in special vacuoles. The vacuole membrane is composed of that of the Golgi vesicles that delivered the building material (Fig. 4, arrowheads).

Fig. 4: On the left, a synoptic representation of the shell of a Paulinella chromatophora, on the right, an optical cross-section through a cell shortly before division. The arrowheads point to the pre-produced shell scales.
Tips for collecting desmids
Distribution and ecology of the species mentioned
Micrasterias rotata is described as acidophilic but adaptable. It prefers a slightly acidic environment (fens and transitional mires), but occurs in acidic sphagnum waters as well as in neutral locations (e.g. meadow wetlands). The tolerated pH range is wide: (3.8)–4.5–7.5! As a characteristic species, it is both common and widespread in Central Europe and is probably cosmopolitan. When it is encountered, it is often found in large numbers.
Micrasterias denticulata is also quite common in fens and transitional mires. If the site conditions suit it, it also tends to multiply in large numbers. Lenzenweger (1996) describes it as a characteristic species for moderately acidic waters of fens and transitional mires, silting and floating grassland zones, and wet meadows and slopes. It is particularly widespread in the area of pre-alpine lowlands, but in the Alps it rises to altitudes of up to 1000 m and is not rare there either.
Micrasterias apiculata has similar ecological requirements, but is rarer.
Micrasterias thomasiana as variety notata is much more widespread than the type variety. Lenzenweger describes it as a characteristic species in acidic to moderately acidic lowland and intermediate moorland loops, wet meadows and forest ponds in the Alpine foothills up to the middle altitudes of the Alps.
As can be read in Coesel & Meesters (2023), the species M. denticulata and M. rotata are rare in the lowland areas of northern Central Europe (M. apiculata very rare), M. thomasiana var. notata is described as “slightly more common”.
Special hints for sampling desmids
The Micrasterias species mentioned above are usually found in very shallow water bodies, in the peat bog pits and depressions in wet meadows. In raised moor areas, it is advisable to look for areas where Sphagnum is no longer the only predominant species, but where there are increasingly vascular plants (grasses) in the water. If you also see Utricularia species (bladderweed) there, you are certainly in the right place, namely in the transitional mire area (John & Williamson, 2009). It is moderately acidic to neutral, and in general you will find the greatest variety of desmid species here.
In shallow and small water areas, the collection method advocated by Lenzenweger using a disposable syringe without needle has proven to be effective. Many desmid species live in the Aufwuchs on Sphangnum species and vascular plants and adhere there with their mucilage for locomotion weakly enough to be detached from the substrate by pulling up the syringe and rubbing the tip back and forth on the growth substrate.
Once the syringe is full, it is advisable to take a closer look through a 10x folding magnifying glass at a few drops that you put on the palm of your hand (Fig. 5). With a little practice, you can even distinguish desmid species groups!

Fig. 5: Sampling with a syringe and checking with a 10x magnifying glass. In the middle section, two examples of peat bog pits in the transitional mire area of the fen “Paradieswiese” near Kitzbühel (Tyrol).
How do you get to see these cell organelles?
Usual state of the algae cells shortly after collection
If you look at freshly collected desmids under a microscope, you will see that they usually contain many spherical assimilate bodies (storage lipids). These include the oil-containing and therefore highly light-refracting plastoglobuli, and the fat-containing oleosomes (Bresinsky & al., 2008). The oleosomes in particular are often so dominant in the image that they obscure the view of delicate and almost translucent organelles such as mitochondria and dictyosomes. Fig. 6 shows some examples:
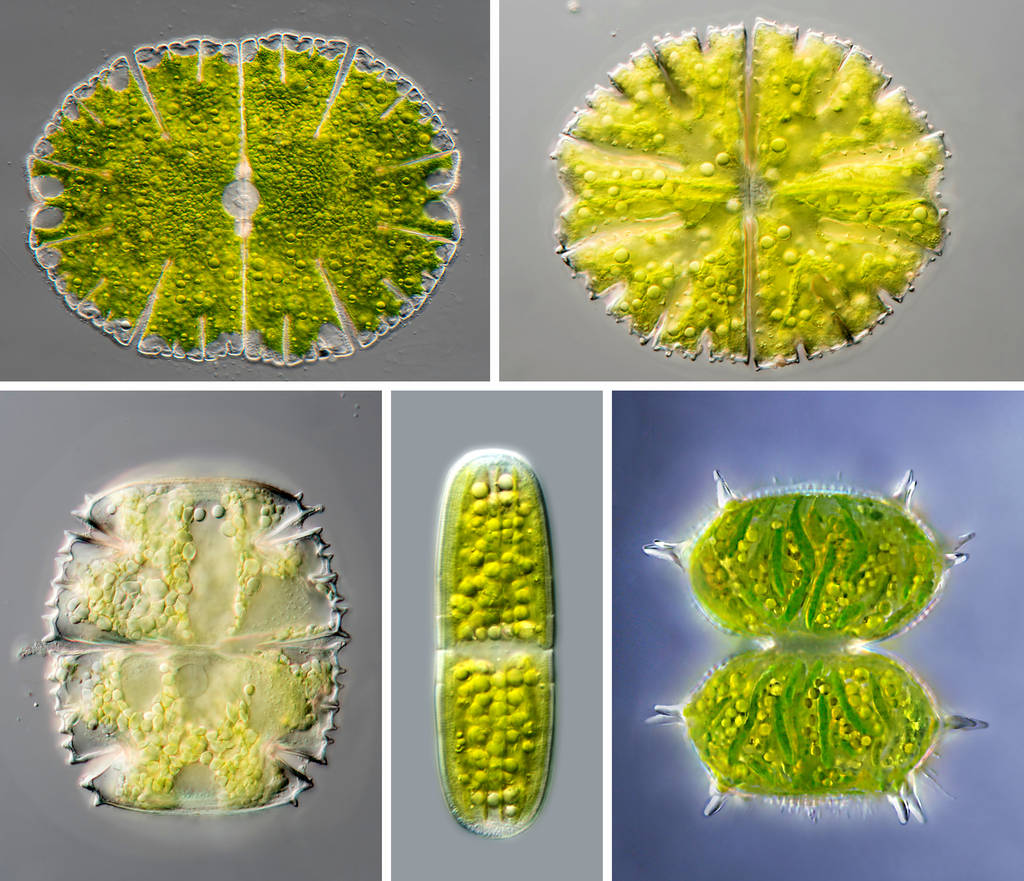
Fig. 6: Assimilate globules in Zygnematophycea cells. Upper row: Micrasterias denticulata and M. papillifera. Lower row: Micrasterias truncata var. bahusiensis, Penium polymorphum and Xanthidium antilopaeum.
The cells must starve
We need to give the cells time to decompose and utilize the fats and oils stored as reserve substances to get a clearer view of the fine cytoplasmic details that we want to observe. Before I could take the images shown below, I had stored my samples in Petri dishes and humid chambers in a basement room for about two months in low light and moderate temperatures (18–20 °C). The fact that the cells in Fig. 2 could be photographed so free of reserve globules is due to such a period of starvation!
Mitochondria and dictyosomes
If the algae are examined at higher magnification (from objective magnifications of 40x with apertures from 1.0), a whole series of structures in the cell plasma can be distinguished. If you slowly focus from the cell surface into the cell lumen, whereby the use of contrast-enhancing measures such as oblique illumination or DIC is helpful, you can see a lively particle stream just below the cell wall. Most of these “microsomes” (membrane-enclosed vesicles of different origins, circled as an example in Fig. 7) are small and round with diameters of less than 0.5 µm and are moved quickly by the cytoplasmic flow in nearly a straight line in areas without obstacles (Kleinig & Sitte, 1984).

Fig. 7: Different particle types in the cytoplasmic stream of Micrasterias rotata. The individual images of the two series of images were taken appr. every second. Arrows = oil drops, arrowheads = mitochondria, circle = vesicle accumulation, “microsomes”. For reasons of clarity, only the movement stages of some mitochondria have been marked. Scale bars indicate 5 µm.
However, in the observations on cells of Micrasterias rotata, the mitochondria proved to be quite location-stable and were rarely displaced, and if they were, then only slowly and only over short distances. They appeared as elongated, shape-changing structures that flapped back and forth in the particle stream like flags in the wind (Fig. 7, arrowheads). Small, highly refractive assimilate spheres (oil droplets) were also repeatedly seen in the particle stream (Fig. 7, arrows).
In M. denticulata, the mitochondria were in constant swinging and dancing motion, just like in M. rotata (Fig. 8, bottom row), but they were more voluminous, the number of mitochondria observed was significantly lower, and they were usually located deeper in the cell, near the large, flat chloroplasts. They were therefore less often photographed as clearly as those in M. rotata. The mitochondria of M. apiculata, on the other hand, were very similar in size and distribution to those in M. rotata (Fig. 8, top row).

Fig. 8, top row: Mitochondria of Micrasterias apiculata together with oil droplets (arrowheads); bottom row: series of images of a mitochondrion of M. denticulata in oscillating motion together with a dictyosome (arrows) and oil droplets (arrowheads). The images of the series were taken at intervals of appr. one second. Scale bars indicate 5 µm.
Dictyosomes are found in cells in much smaller numbers than mitochondria. Their shape appears round and flat under the light microscope, and their surface and edge appear to be nubbed. Their diameter is usually 0.5–2 µm, in rare cases around 5 µm (in von Denffer & al., 1971, there is an explicit reference to Micrasterias!). Similar to mitochondria, they are usually not displaced by the plasma flow; if they are, they swing back again and thus remain in a certain area. They are concentrated in the area around the nucleus and the edges of the chloroplast wings (Fig. 8, bottom row and 9). Tilting movements were also rarely seen, but in one case a few profile images were obtained (Fig. 9, top row).

Fig. 9: Serial photographs of dictyosomes from Micrasterias denticulata (top row) and M. thomasiana var. notata (bottom row). The typical tilting movements mentioned in the literature (Drawert & Mix, 1962) are clearly visible, especially in the first row (images 3 and 4) . Scale bars indicate 5 µm.
The mobility of the desmids
Many desmid species can move in a targeted manner using mucilage (Linne von Berg & al., 2003). They have the ability to align themselves with the light and move in the direction of the light, even perpendicular to the substrate! When it gets dark, Micrasterias cells position themselves vertically. This gives them the largest possible “sensor surface” for the expected direction of light at the start of the next period of light. At dawn, they orient their chloroplasts perpendicular to the light. The scientific film of IWF C1496 by Wenderoth (1983) shows this impressively.

Fig. 10, image on the left: Mucilage produced for locomotion purposes on a cell tip of Closterium moniliferum; image on the right: After a dark phase, a cell of Micrasterias rotata attached vertically to the slide using mucilage, photographed with an inverted microscope. Scale bar indicates 100 µm.
They have an additional, specialized type of dictyosomes for the production of the mucilage for locomotion, which consists of polysaccharides. When you see the voluminous expulsion of mucilage for locomotion purposes in the film mentioned, you can’t help but wonder where the cells get this abundance of material from. The process can only be understood if you assume a high swelling rate. In fact, it was measured at the root tips of corn, for example, that the mucilage exuding as a lubricant was 1000 times more voluminous than the internal supply of mucilage vesicles to the outflow openings of the root tips (Steer, 1985).
Locomotor mucilage-producing dictyosomes
When the storage lipids have been broken down and consumed, a look at Micrasterias rotata reveals a few round structures spread over the large cell surface, the size of which is similar to that of the large lipid spheres that dominate the picture of freshly collected cells. On closer inspection, however, one notices clear differences; unlike the storage substances mentioned, most of them do not lie motionless, they oscillate slightly around an imaginary center. Fig. 11 shows an overview image.
In Fig. 12, the cytoplasmic bodies in question are shown side by side in two photomicrographs of Micrasterias apiculata. The algae sample from which this cell came had not been placed in twilight for quite as long, so some of the larger reserve substance spheres were still in the cytoplasm. The latter have a high optical density and are also easy to see in bright field, while the delicate dictyosomes are usually only clearly visible with the help of optical contrast methods such as oblique lighting or DIC. Fig. 13 shows impressive examples at maximum magnification for light microscopy.

Fig. 11: Micrasterias rotata after two months in a humid chamber set up in a poorly lit room. The round bodies are the dictyosomes, which are specialized in the production of mucilage for locomotion purposes. Scale bar indicates 50 µm.

Fig. 12: Micrasterias apiculata, sections close to the cell nucleus with dictyosomes (arrowheads), highly refractive plastoglobuli (double headed arrow) and oleosomes (arrows). Scale bar indicates 5 µm.

Fig. 13: Mucilage-producing dictyosomes of Micrasterias rotata, in the first image together with an oil droplet. In the third image, a pattern is visible that indicates vesicle structures. The dictyosome in the fourth image was positioned so favorably that a brightfield image was possible without the aid of oblique lighting. Scale bar indicates 5 µm.
Vesicle detachments
In her electron-optical studies of dictyosomes of Micrasterias americana that produce motion mucilage, Noguchi (1978) showed that huge vesicles with diameters of 1.2 to 1.8 µm are formed. I was able to observe such vesicle formation and detachment several times; it took an average of about four minutes from the first noticeable bulge at the edge to the detachment (Fig. 14). An early stage of such vesicle formation can also be seen in the short film sequence I recorded below.

Fig. 14: Micrasterias rotata. Detachment of a large vesicle from a dictyosome. Scale bar indicates 5 µm.
A short video showing locomotive mucilage-producing dictyosomes in
Micrasterias rotata along with “Microsomes” (vesicles of different origins) and oil droplets.
From about the position “second 25” the starting phase of a detachment of a Golgi vesicle becomes visible at the edge of the centrally located dictyosome at 5 o’clock.
Concluding remark
Electron-optical images show that dictyosomes are generally surrounded by a ring of small membrane-enclosed vesicles, the Golgi vesicles. The smooth-edged objects in the above images do not initially match the expected image. However, due to the limited resolution of the light microscope, the small and smallest constrictions cannot be shown. If the dictyosomes were close to the cover glass, sometimes at least faint outlines of vesicles could be documented (Fig. 13, third individual image). Even after switching to bright field, some dictyosomes remained visible if they were in a favorable position (Fig. 13, fourth individual image). One dictyosome showed a constriction in the central area (Fig. 13, second individual image). Electron-microscopic images were found in the literature (Menge & Kiermayer, 1977; Noguchi, 1978) which indicate that this image could correspond to a division stage.
It should also be noted that all photomicrographs of small and tiny cell organelles shown here were taken on intact algal cells.
Acknowledgements
I would like to express my sincere thanks to Dr. Detlef Kramer, cytologist and at the time head of the electron microscopy laboratory at the Technical University of Darmstadt (my photomicrographs of cell biological details are from 2007), for his friendship, encouragement, valuable discussions and joint microscopic investigations on the topic. I am also very grateful to Prof. Dr. Werner Herdt, who was a cell biologist at the University of Heidelberg at the time, with whom I was able to discuss the findings I had obtained using light microscopy and who, following our discussions, verified these findings by conducting his own electron microscopic studies on Micrasterias rotata.
References
Bresinsky, A., Körner, C., Kadereit, J. W., Neuhaus, G., & Sonnewald, U. (2008). Strasburger, Lehrbuch der Botanik für Hochschulen. 36th edition/revised by Andreas Bresinsky. Spektrum, Akad. Verl. Heidelberg.
Coesel, P. F., & Meesters, K. J. (2023). Desmids of the Lowlands: Mesotaeniaceae and Desmidiaceae of the European Lowlands. 2nd revised and expanded edition: 424 pages, 26 figures and 3 tables in the text and 136 plates. KNNV Publishing, Zeist, the Netherlands.
Drawert, H., & Mix, M. (1962). Zur Frage von Struktur und Funktion des „Golgi-Apparates“ in Pflanzenzellen. Marb.Sitzungsber., 84 (1. H), 361–382.
Herth, W. (2007). Personal communications.
Hess, S., Williams, S. K., Busch, A., Irisarri, I., Delwiche, C. F., de Vries, S., Darienko, T., Roger, A. T., Archibald, J. M., Buschmann, H., v. Schwartzenberg, K. & de Vries, J. (2022). A phylogenomically informed five-order system for the closest relatives of land plants. Current Biology, 32(20): 4473–4482.
John, D. M., & Williamson, D. B. (2009). A Practical Guide to the Desmids of the West of Ireland: VIII, 196 pages, 117 figures in the text and 49 plates. Martin Ryan Inst., National University of Ireland, Galway.
Kleinig, H., & Sitte, P. (1984). Zellbiologie: XI, 488 pages, 482 figures and 87 tables.. Gustav Fischer Verlag, Stuttgart.
Kramer, D. (2007). Personal communications.
Lenzenweger, R. (1996). Desmidiaceenflora von Österreich. Bibliotheca Phycologica. B 101: VIII, 162 pages, 117 figures in the text and 17 plates. J. Cramer in der Gebrüder Borntraeger Verlagsbuchhandlung, Berlin – Stuttgart.
Linne von Berg, K.-H., Hoef-Emden, K., Marin, B., Melkonian, M. (2003). Der Kosmos-Algenführer: 366 pages, 199 graphics in the text, 202 graphics in the key and 783 color photos. Franckh-Kosmos Verlag, Stuttgart.
Menge, U. & Kiermayer, O. (1977). Dictyosomen von Micrasterias denticulata Breb. – ihre Größenveränderung während des Zellzyklus. Protoplasma 91, 115–123.
Noguchi, T. (1978). Transformation of the Golgi Apparatus in the Cell Cycle, especially at the Resting and Earliest Developmental Stages of a Green Alga, Micrasterias americana. Protoplasma 95, 73–88.
Ružička, J. (1981). Die Desmidiaceen Mitteleuropas, Volume 1, part 2: IX, 293–736, 4 illustrations in the text and 73 plates. E. Schweizerbart’sche Verlagsbuchhandlung (Nägele u. Obermiller), Stuttgart.
Steer, M. W. (1985): Vesicle dynamics. In Robards, A., W. (ed): Botanical Microscopy 1985. Oxford University Press: 129–155.
v. Denffer, D., Schumacher, W., Mägdefrau, K. & Ehrendorfer, F. (1971). Lehrbuch der Botanik für Hochschulen „Strasburger”. 30th edition: VIII, 842 pages, 759 illustrations in the text and 1 colour map. Gustav Fischer Verlag, Stuttgart.
Wenderoth, K, & Inst. Wiss. Film (1983). Phototaxis bei Desmidiaceen und Diatomeen. Film C1496 of the IWF, Göttingen.
Wenderoth, K. (1985). Phototaxis bei Desmidiaceen und Diatomeen. Publ. Wiss. Film., Sekt. Biol., Ser. 17, Nr. 15/C1496.
© Wolfgang Bettighofer,
images under Creative Commons License V 3.0 (CC BY-NC-SA).
For permission to use of (high resolution) images please contact postmaster@protisten.de.
New items

elegantissimiforme
pseudowillei
antilopaeum
laeve
basiornatum
capitulum
spec.
africana
caelatum


pinnatifida

striolatum
decedens

acutiformis
astraea

bipyramidatum

cinctum
pulchrum

melicerta
lunatus

brebissonii
montanum

directum

truncatum

vestitum
var. vestitum
meneghinianum
didelta

furcata

jenneri
ventricosum
insigne
cucurbita
fimbriata

cucurbita

wailesii
filifera

brightwellii
New items
elegantissimiforme
pseudowillei
antilopaeum
laeve
basiornatum
capitulum
spec.
africana
caelatum
pinnatifida
striolatum
decedens
acutiformis
astraea
bipyramidatum
cinctum
pulchrum
melicerta
lunatus
brebissonii
montanum
directum
truncatum
vestitum
var. vestitum
meneghinianum
didelta
furcata
jenneri
ventricosum
insigne
cucurbita
fimbriata
cucurbita
wailesii
filifera
brightwellii